

ウナギの回遊研究と資源問題
東京大学海洋研究所 青山 潤・塚本勝巳
1.はじめに
ウナギは、我が国で年間15万トンも消費される重要な水産資源である。夏の盛り、町に流れる香ばしい蒲焼きの匂いは、ひとつの風物詩となっている。さらに、ウナギは古くから民話や信仰の対象として、日本の文化と深く関わってきた。すなわち、日本人にとって、ウナギは最もなじみ深い魚の一つであるといえる。
一般に川の魚と考えられがちなウナギ Anguilla japonicaであるが、実は、およそ3000キロ離れたグアム島付近の海で産卵する「降河回遊魚」である。外洋でふ化したウナギは、透明な柳の葉のような形をした仔魚(レプトセファルス)として、海流に流されながら東アジア一帯へ来遊する。そこで、親と同じ形をした透明なシラスウナギへ姿を変え、沿岸・河口域へ接岸する。私たちが普段口にする蒲焼きは、ほぼ百パーセントこうして捕らえたシラスウナギを、養殖池で大きくしたものである。
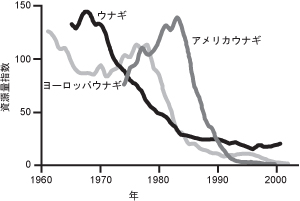
しかしながら、シラスウナギの漁獲量は1970年代をピークに確実に減少を続け、現在では最盛期の10-20%程度で推移している(図1)。一方、大西洋にもアメリカウナギ A. rostrataとヨーロッパウナギ A. anguillaが生息し、特にヨーロッパの一部では重要な水産資源の一つとなっている。興味深いことに、大西洋のウナギ資源も、太平洋のウナギにややおくれ70~80年代から大きく減少しているのである。さらに近年、ヨーロッパウナギの資源水準は盛時の1%以下にまで減少し、レッドデータブックへの記載が検討されるまでになった。
2.ウナギの産卵場
ウナギA. japonicaの産卵場はマリアナ諸島西方海域とされている1)。しかしながら、厳密な意味では、ウナギの産卵場の位置はまだ特定されていない。これは世界中のどのウナギ属魚類についても同様で、ウナギの産卵現場のみならず天然の卵すら見た者はいないのである。
ウナギの産卵場調査は1930年代に始まったが、最初のレプトセファルスが採集されたのは、1967年になってからである。その後、計4回の大規模な研究航海の末、1991年7月、ついに東京大学海洋研究所の白鳳丸(現在は海洋研究開発機構所属)が全長10 mm前後のウナギのレプトセファルス約1000尾を採集し、マリアナ諸島西方海域が産卵場であることを明らかにした1)。研究はさらに継続され、それまでに得られた全てのレプトセファルスの分布、体サイズ、海流や海底地形から、ウナギの産卵場は西マリアナ海嶺北緯15°前後の3つの海山(スルガ、アラカネ、パスファインダー)であると推定された2、 3)。また、採集されたレプトセファルスの孵化日解析から、ウナギは産卵期各月の新月の日に同期して一斉に産卵することがわかった2)。ウナギの産卵場と推定された海山は、水深3000~4000mの海底からほとんど海表面近く(水深10~40m)までそびえ立つ富士山(3776m)クラスの高い山々である。これらの海山は東アジアから3千キロの大回遊をしてきたウナギの雌雄が集合する出合いの場であり、例えば新月の翌日に産卵場へ辿りついたウナギが産卵のため次の新月の日まで待機する待合所でもある。さらにこれら海山が含まれる西マリアナ海嶺は昔火山であったことからここには磁気異常が生じ、東アジアから南下して産卵に向かう親ウナギの道しるべとなっている可能性もある。
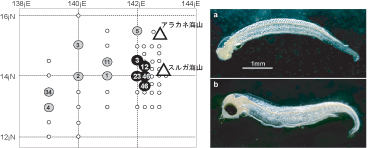
これら2つの仮説により、世界のウナギ属魚類で初めて産卵の時間と場所を厳密に絞り込むことができた。そして、これに基づき実施された2005年6月の白鳳丸による研究航海の結果、新月である7日に産卵場と推定されていたスルガ海山の西約100kmの地点で、孵化後2日齢のプレレプトセファルス約400尾が採集された(図2)4)。海流の平均流速と日齢および卵期の長さから逆算するとこれらのプレレプトセファルスは、新月の4日前にスルガ海山近傍で産卵されたものであることがわかった。これらのことはウナギ産卵場に関する上記2つの仮説を満たしており、ウナギの産卵地点をピンポイントで突きとめたといえる。これにより30余年におよぶウナギ産卵場調査はひとまず終止符を打つことになった。
ウナギの産卵現場やその繁殖生態を明らかにすることは、ウナギの人工催熟や仔魚飼育技術に重要な示唆を与え、人工種苗生産技術の飛躍的な向上に寄与する。さらに今後、ウナギ卵や親魚が採集されれば、ウナギの生物学における大きな進展となり、産卵生態・繁殖生理学などの研究は全く新しい境地へ入ることとなる。
3.レプトセファルスの回遊
産卵場で生まれた卵は、プレレプトセファルス、レプトセファルスを経て、シラスウナギへと変態し、東アジア一帯の沿岸・河口域へ接岸してくる(図3)。この間の初期生活史は、耳石日周輪解析や波長分散型X線分析装置によるストロンチウム濃度の分析からかなり明らかにされている。
まず、シラスウナギ5)、孵化仔魚6)さらにレプトセファルス7)を用いて耳石輪紋形成の日周性が確認された。また、ウナギの耳石には、ストロンチウム/カルシウム比の急減と同期した耳石輪幅の急増点が存在し、これがレプトセファルスからシラスウナギへの変態開始を示す印であることもわかった。これらの技術を用いて、東アジア各国へ接岸したシラスウナギの回遊生態は、おおよそ明らかにされている。すなわち、レプトセファルスは1日に0.5mmずつ成長しながら北赤道海流で西へ輸送され、やがて黒潮に乗換え北上する。また、孵化後150日前後で全長約60mmに到達すると変態を開始し、約3週間かかってシラスウナギになることもわかった8)。すなわち、黒潮を離脱したシラスウナギは東アジアの河口を目指し、孵化後180日前後で河口域に到着するのである。さらに、接岸回遊の際、産卵期の早期に孵化したものほど、若齢で、早期に東アジアの低緯度域に加入する傾向が認められている8)。
一方、レプトセファルスが来た赤道海流から北上する黒潮へ乗り換える際に貿易風が重要な役割を果たすことがわかってきた(貿易風仮説 trade wind Hypothesis)9)。ウナギが首尾よく東アジアにやって来るには適度に貿易風が吹くことが重要で、強すぎても、弱すぎてもいけないというのである。強すぎる場合は黒潮へ取り込まれる前に北上しすぎて黒潮反流の渦の中にとりこまれてしまい、逆に弱すぎた場合は、南下するミンダナオ海流に取り込まれ、本来ウナギの分布しないフィリピン南部やインドネシアへ輸送され、無効分散(死滅回遊)になってしまう。事実、無効分散の例を示す全長40mm以上のかなり発育したウナギのレプトセファルスがセレベス海で採集されている(Miller et al. 未発表)。この貿易風仮説はシラスウナギの接岸量を予測する唯一の仮説として注目されている。また、エルニーニョによって産卵場の塩分フロントが南北に移動することでレプトセファルスの輸送環境が大きく変化し、シラスウナギの接岸量が変わるという「エルニーニョ仮説 El Nino Hypothesis」も提出されている。 今後、これらレプトセファルスの回遊生態に関するメカニズムが明らかにされれば、シラスウナギの接岸量の予測や、ウナギ資源の管理・保全方策の立案に重要な示唆を与えることは間違いない。
4.海ウナギ
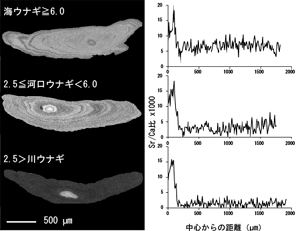
波長分散型X線分析装置を用いて、耳石に含まれる微量元素のストロンチウム分布を調べることにより、その魚が生活史のどの時期に海にいて、いつ淡水に入ったかを推定することができる。すなわち、個体の回遊履歴がわかる。これは海水中のストロンチウム濃度が淡水中のそれより高く、魚類は環境水中のストロンチウムを体内に取り込み、耳石の成長層を形成するからである。
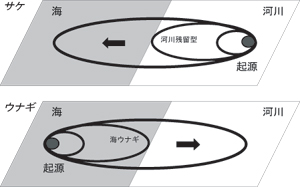
日本沿岸で採集された産卵回遊中の銀ウナギ283尾の耳石微量元素分析を行ったところ、降河回遊魚に分類されるウナギの中に、河川に遡上せず、一生海で過ごす"海ウナギ"の多数いることが発見された(図4)10)。これは、遡河回遊魚のサケ科魚類における河川残留型に対応する非回遊型の個体群と考えられている(図5)。海ウナギの存在は、ウナギの回遊行動の進化における"先祖返り"とも解釈できる生活史多型である11)。また、高緯度へいくほど、海ウナギの出現率が高くなる傾向が認められ(Kotake et al. unpublished)、サケ科魚類のスモルト降海率と逆の緯度クラインをもつものと考えられた。
通常の降河回遊型の"川ウナギ"は日本沿岸の銀ウナギ全体に占める割合が14.5%と低く、逆に淡水を全く経験しない"海ウナギ"や"河口ウナギ"が計85.5%もの高率を示すことが明らかにになった。このことは、本種のマリアナ沖の産卵場における再生産が現在のところ、主にこれら非回遊型の個体群によって支えられていることを示唆している10)。
海ウナギの発見はウナギの生活史に多型があり、その回遊が可塑性に富んだものであることを示すだけでなく、魚類の回遊行動の進化過程を考察する上でも重要であると考えられる。さらに海ウナギに関する知見は増殖、保全対策立案にも重要な基礎資料となる。
5.終わりに
ここに上げたのは、資源の管理・保全方策の立案に根本的な影響を与えるいくつかのウナギの生態学的特徴に過ぎない。今後は何よりもまず、未だ多くの謎に包まれているウナギの生活史の全貌を明らかにする必要がある。また先に述べたように、同一の産卵集団であるにもかかわらず、ウナギ個体群の量や性比、サイズ組成、成熟年齢などは成育場によって大きく異なる。すなわち、ウナギ資源の実態を明らかにし、適切な管理・保全を行うためには、日本、台湾、韓国、中国などウナギ資源を共有する東アジア諸国の協力が必要不可欠となる。事実、1998年には研究者や関連業界団体からなる「東アジアウナギ資源協議会」(East Asia Eel Resource Consortium: EASEC)が設立され、毎年一回すでに計8回の国際会議を開いている。しかしこの会合では、毎回ウナギ資源に関する興味深い研究報告と議論が行われているものの、管理・保全の主体となる行政の参加がなく、実際の保全対策に会議の成果が直接生きてこない点がやや不満である。ウナギ資源の生物学的特性を理解し、「我が国のウナギ」ではなく東アジア全体の共通財産として、最大消費国の日本が強いリーダーシップをとることにより、ウナギ資源の管理・保全対策を早急に実施することが望まれる。
参考文献
- Tsukamoto K. Discovery of the spawning area for the Japanese eel. Nature. 1992; 356: 789-791.
- Tsukamoto K, Otake T, Mochioka N, Lee T-W, Fricke H, Inagaki T, Aoyama J, Ishikawa S, Kimura S, Miller MJ, Hasumoto H, Oya M, Suzuki Y. Seamounts, new moon and eel spawning: the search for the spawning site of the Japanese eel. Environ. Biol. Fish. 2003; 66: 221-229.
- Tsukamoto K, Fricke H, Lee T-W. Spawning area of the Japanese eel. In: Aida K, Tsukamoto K, Yamauchi K (eds) Eel biology. Springer, Tokyo. 2003; 121-140.
- Tsukamoto K. Spawning of eels near a seamount. Nature, 2006; 439: 929.
- Tsukamoto K. Otolith daily increments in the Japanese eel. Bull. Jpn. Soc. Sci. Fish. 1989; 55: 1017-1021.
- Umezawa A, Tsukamoto K, Tabeta O, Yamakawa H. Daily growth increments in the larval otolith of the Japanese eel, Anguilla japonica. J. Ichthyol. 1989; 35: 440-444.
- Shinoda A, Tanaka H, Kagawa H, Ohta H, Tsukamoto K. Otolith microstructural analysis of reared larvae of the Japanese eel Anguilla japonica. Fish. Sci. 2004; 70: 339-341
- Tsukamoto K. Recruitment mechanism of the eel, Anguilla japonica, to the Japanese coast. J. Fish Biol. 1990; 36: 659-671
- Kimura S, Tsukamoto K, Sugimoto T. A model for the larval migration of the Japanese eel: roles of the trade winds and salinity front. Mar. Biol. 1994; 119: 185-190.
- Tsukamoto K, Nakai I, Tesch FW. Do all freshwater eels migrate? Nature. 1998; 396: 635-636.
- Tsukamoto K, Aoyama J, Miller MJ. Migration, speciation and the evolution of diadromy in anguillid eels. Can. J. Fish. Aquat. Sci. 2002; 59: 1989-1998.
*図1
世界のウナギ資源の動向
*図2
2005年6月7日、白鳳丸によって孵化したてのウナギ仔魚が採集されたマリアナ沖の測点(左パネル)。黒丸は6月生まれのプレレプトセファルス、白丸は5月生まれのレプトセファルスの採集測点。スルガ海山のすぐ西でプレレプトセファルスが採集された。右パネルは、新月の日に採集された2日齢のプレレプトセファルス(上). 眼も歯もまだ形成されていない. 下はその3日後に採集された5日齢の個体。
*図3
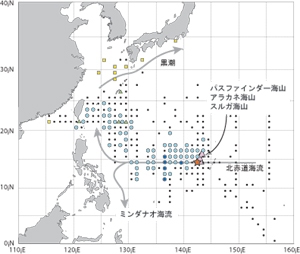
外洋におけるウナギ仔稚魚の分布。1956-2002年までに行われた31回の研究航海で採集された計2418個体の採集測点を示す。●はレプトセファルス、▲は変態仔魚、■は海洋シラスの採集測点をそれぞれ示す。記号の大きさはその測点の採集個体数に対応する(小:1個体、中:2-9個体、大10個体以上)。小黒点は採集努力を払ったにもかかわらず、ウナギの仔稚魚が採集されなかった測点を示す。海山仮説の3つの海山(大きな三角印:スルガ海山、アラカネ海山、パスファインダー海山)はおよそ北緯15°東経142.5°にある。図中星印は孵化後2日齢の孵化仔魚(プレレプトセファルス)の採集地点を示す。
*図4
日本沿岸域で採集された産卵回遊中の銀ウナギの耳石Srの分布(左)と、EPMAによる線分析結果(右、Kotake et al., unpublished)。耳石中心部はレプトセファルス期に海洋中で形成されるのでSr濃度は高い値を示し(中心部の明部)、シラスウナギへの変態に伴って急減する。その後、上段の海ウナギではSr/Ca比が7前後の高値を維持して、終生海域で過ごしたことを示すのに対し、下段の川ウナギは、2前後以下の値を示し、長い淡水生活期を表している。中段の河口ウナギはこれらの中間的な値を示す。
*図5
サケとウナギの通し回遊の起源と進化11)。 種の産卵場と成育場を結ぶ概念的回遊経路を回遊環(Migration loop)と名付け、回遊環の変化より回遊行動の進化を論じた。